
メンバー紹介

教授:市川 進一
最終出身校
1984年3月 東京薬科大学薬学部卒業
1986年3月 東京工業大学大学院修士課程修了
学位
博士(理学)(東京工業大学1992年2月)
職歴
ペンシルバニア大学医学部Research Specialist(1986年~1989年)
北里大学薬学部 微生物薬品製造学 講座研究員(1989年~1990)
名古屋大学医学部 生化学第一講座 助手(1990年~1992年)
理化学研究所 フロンティア研究員(1992年~2000年)
(株)エイジーン研究所 主任研究員(2000年~2001年)
(株)ジーンケア研究所 創薬部 部長(2001年~2002年)
新潟薬科大学 応用生命科学部 助教授(2002年~2007年)同教授(2007年~)
資格
薬剤師
受賞
第57回注目発明(科学技術庁)
研究分野
分子生物学 生化学 神経化学 糖鎖生物学
(1) スフィンゴ脂質(特にグルコシルセラミド)の研究
(2) DNA傷害、細胞ストレスによる細胞死の研究(特に老化とがんに関係する部分)
(3) 脂肪肝の抑制に関する研究
(4) 痛みの研究
学部生
修士課程2年生
斎藤 泉
修士課程1年生
関根 大晃
長谷川 雄太
西川 瑞穂
鈴木 聡珠
学部4年生
伊部 萌水
遠藤 彩音
佐波 春乃
高橋 絵里子
佐藤 哲哉
卒業生の進路
卒業生の進路
研究室開設以来12期の学生が卒業して行きましたが、就職については就職を希望した学生のほとんどが無事就職しています(12年間の平均は、進学を希望しなかった学生も含めて学部卒が97%、大学院卒が100%です)。就職先は以下の通りです。
卒業生の就職先
学部:
ドクターシーラボ、中外製薬、デンカ生研、バイオテックジャパン、
星光堂薬局、クスリのアオキ、ブルボン、亀田製菓、たいまつ食品、
安田ヨーグルト、日東アリマン、セイヒョー、フタバ、宮野食品工業所、
武蔵野、東北日本ハム、アーデン、NHC、大光銀行、ゆうちょ銀行、
かんぽ生命保険、丸福証券、アース環境サービス、ナルス、ウオロク、
新潟酒販、アークランドサカモト、シバタ医理科、共立医科器械、
サンカ、JAえちご上越、パラッツォ、新潟サンリン、ミカサ、
キューピット、THK新潟、VIP、テイク・フォーなど。
大学院:(本研究室卒業生のみ):
アース環境サービス、日本臓器製薬、亀田製菓、鍋林、三幸、
WDBエウレカ、陽進堂、和光純薬工業
卒業生の大学院進学先
弘前大学、東北大学、東京大学、
東京工業大学、東洋大学、
横浜市立大学、筑波大学、群馬大学、
新潟大学、新潟薬科大学、
奈良先端科学技術大学院大学、
名古屋大学、神戸大学、京都大学、
九州大学
研究テーマ
1.スフィンゴ脂質の研究
スフィンゴ脂質は、細胞膜上に存在し、多数の分子種から成る一群の物質です。最も基本的なスフィンゴ脂質はセラミドで、この物質に様々な種類の糖鎖が付加されて、スフィンゴ糖脂質となります。スフィンゴ糖脂質は糖鎖の違いにより多くの分子種が知られており、細胞間相互作用、分化誘導、細胞接着など、様々な現象に関係しています。スフィンゴ脂質の代謝異常は、ゴーシェ病、糖尿病、多剤耐性がんなど様々な病気の原因になります。スフィンゴ糖脂質の合成は、グルコシルセラミド合成酵素による、セラミドのグルコシル化反応から始まります。一方で、グルコシルセラミド合成酵素の基質であるセラミドは、細胞を自殺に導くシグナル分子であると考えられるようになってきています。このため、グルコシルセラミド合成酵素の活性上昇がセラミドの量を減少させ細胞死を抑制していると考えられるようになってきました。私たちはグルコシルセラミド合成酵素の遺伝子の発現や酵素活性の変化を調べることにより、どのような時にグルコシルセラミド合成酵素が細胞死を抑えるかを調べています。また、グルコシルセラミド合成酵素を持たない細胞やその阻害剤を使って、スフィンゴ糖脂質の体の中での働きを調べています。今後、グルコシルセラミドを中心とするスフィンゴ脂質類の今まで知られていない生体内の働きを見つけて行きたいと考えています。
最近は、医薬品や機能性を持つ食品の開発を目指して、グルコシルセラミド合成酵素を阻害する化合物、食品および植物の探索も行っています。

グルコシルセラミド合成酵素活性の検出
2.細胞死を抑制する遺伝子の研究
ヒトの体を構成する生体成分は、常に外部からの放射線、化学物質や、体内で代謝により生成する毒性物質、活性酸素などにより損傷を受け続けています。生体には、異常が起きた物質や細胞を排除し、再生産する機構が備わっていますが、その働きは完全ではありません。異常な生体成分の蓄積、特に遺伝子に蓄積される変異は、ガンを含む多くの疾患や老化の原因となっています。近年、分子生物学の進歩により、多くの種類のガンが、複数のガン遺伝子やガン抑制遺伝子の変異によって生じることが明らかにされてきました。一方で種々の早老症(老化が正常人よりも早く進む疾患)の研究から、遺伝子の修復や、ゲノムの安定化に関係する、老化抑制遺伝子の存在が明らかになってきています。これらの遺伝子の研究から、老化の主要な原因は、遺伝子に蓄積した異常や細胞にかかるストレスによる細胞の機能不全と、それに続いておきる細胞死による、細胞の減少だと考えられるようになってきています。
そこで、私たちは活性酸素やDNA傷害によって起きる細胞死を抑制する遺伝子群の探索を試みています。方法としては発現クローニングという方法を用いています(図)。まず、培養細胞にレトロウィルスベクターを用いて7.5日胚由来の cDNA ライブラリーを導入します。マウスの7.5日胚は、ちょうど体の形ができ始める時期にあたり、多種多様な遺伝子が発現しています。また、一般的に発生の初期ほど遺伝子の防御、修復能力が高いと予測されています。個々の細胞にはcDNAライブラリーから、それぞれ異なったcDNAが導入され染色体DNAの中に組み込まれます。また、そこから作られるタンパク質によって各々が異なった性質を持つようになります。次に、このような細胞の集団を、DNA傷害性薬剤や活性酸素を発生する薬剤で処理し細胞死を誘導します。目的のcDNAを導入された細胞は薬剤に対して抵抗性となり生き残るので、これらの細胞から導入されたcDNAを再び取り出して分析します。現在、活性酸素の一つである過酸化水素については、細胞死を抑えるcDNA(遺伝子)を数種類見つけており、その作用機構について分析を行っています。これら遺伝子を調べることにより細胞死を抑制する経路について新たな知識が得られ、将来老化やがんの研究につながるのではないか期待しています。
3.脂肪肝の研究
食べ物を食べすぎると、エネルギーとして使われなかった栄養の一部は、中性脂肪となって肝臓の細胞にたくわえられます。こういった中性脂肪は細胞の中で脂の粒(脂肪滴)になります。このようにして肝臓の細胞の中で脂肪滴を持つ細胞が多くなった状態を脂肪肝(非アルコール性脂肪肝)とよびます。フランス料理に使われるフォアグラは、人間がトリに餌を過剰に食べさせて作った脂肪肝です。食べ過ぎによって起きる脂肪肝は今まであまり体に害がないと考えられていました。しかし、最近脂肪肝の一部が、炎症を起こし肝炎(非アルコール性脂肪肝炎)になり最終的に肝臓がんの原因になることが分かってきています。私たちはシャーレの中で培養した肝臓細胞株を用いて、脂肪肝を抑える食品、植物や化合物の探索を行っています。また、脂肪肝を抑えることが分かった物については、その仕組みを生化学的手法や分子生物学的手法を用いて分析しています。
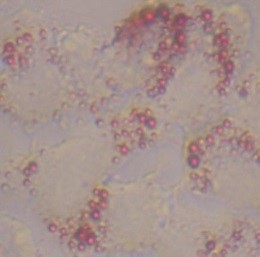
培養細胞の脂肪肝モデル 赤い粒子が脂肪滴
4.痛みの研究
けがなどをした時に、長い間強い痛みを感じていると、傷がなおったあとも痛みが消えないことがあります。こういった痛みを慢性痛といいます。私達はある特定の種類の慢性痛の形成に特定のイオンチャネルの異常が関係しているのではないかと考えて研究を行っています。また、細胞膜上の脂質の組成が各種イオンチャネルに及ぼす影響についても調べています。

イオンチャネル活性の測定
研究成果
- Tsurumaki, H. Katano, H., Sato, K., Imai, R., Niino, S., Hirabayashi, Y. and Ichikawa, S. (2017) WP1066, established small molecule inhibitor of Janus kinase/STAT3 pathway inhibits ceramide glucosyltransferase activity Biochem. Biophys. Res. Commun. 491, 265-270
- Takahash, T., Sugawara, W., Takiguchi, Y., Takizawa, K. Nakabayashi, Y., Nakamura, Y., Nagano-Ito, M. and Ichikawa, S. (2016) Identification of plants that inhibit lipid droplet formation in liver cells: Rubus suavissimus leaf extract protects mice from high fat diet-induced fatty liver by directly affecting liver cells Evid. Based. Complement. Alternat. Med.(eCAM) 2016, Article ID 4282758, 9 pages
- Niino, S., Nakamura, Y., Hirabayashi, Y., Nagano-Ito, M. and Ichikawa, S. (2013) A small molecule inhibitor of Bcl-2, HA14-1, also inhibits ceramide glucosyltransferase Biochem. Biophys. Res. Commun. 433, 170-174
- Nagano-Ito, M and Ichikawa, S (2012) Biological effect of mammalian Translationally Controlled Tumor Protein (TCTP) on cell death, proliferation, and tumorigenesis Biochem. Res. Int. 2012, Article ID 204960, 7pages
- Aida, J., Higuchi, S., Hasegawa, Y., Nagano-Ito, M., Hirabayashi, Y., Banba, A., Shimizu, T., Kikuchi, A., Saga, M., and Ichikawa, S. (2011) Up-regulation of ceramide glucosyltransferase during the differentiation of U937 cells J. Biochem. 150, 303-310
- Nagano-Ito, M., Banba, A., and Ichikawa, S. (2009) Identification of genes that suppress oxidative-stress induced cell death by functional cloning: TCTP prevents hydrogen peroxide-induced cell death FEBS lett. 583, 1363-1367
- Nagano-Ito, M., Tamura, M., Tsurumaki, M., Yoshikawa, S., and Ichikawa, S. (2007) Identification and characterization of a novel alternative splice variant of mouse GMx33a/GPP34 Gene 400, 82-88
- Miyoshi, I., Okamura,T., Kasai, N., Kimoto, T., Ichikawa, S., Osuka, S., Hirabayashi, Y. (2006) Conventional/conditional knockout mice; Sphingolipid Biology (Eds, Hirabayashi Y., Igarashi, Y., and Merrill, A. H., Springer) pp.443-451
- Uchida,Y., Itoh,. Taguchi,Y., Yamaoka, S., Umehara1,H., Ichikawa,S., Hirabayashi Y., Holleran,W.M. and Okazaki, T. (2004) Ceramide reduction and transcriptional up-regulation of glucosylceramide synthase through Doxorubicin-activated Sp1 in drug-resistant HL-60/ADR cells Cancer Res. 64, 6271-6279
- Stanic, A. K., De Silva. A. D., Park, J-J., Sriram, V., Ichikawa, S., Hirabayashi Y., Hayakawa, K., Kaer, L. V., Brutkiewicz, R. R., and Sebastian, J. (2003) Defective presentation of the CD1d-restricted natural Va14Ja18NKT lymphocyte antigen caused by b-D-glucosylceramide synthase deficiency Proc. Natl. Acad. Sci. USA 100, 1849-1854
- Uchida, Y. Murata, S., Schumuth, M., Behne, M. J., Lee, J. D., Ichikawa, S., Elias, P. M., Hirabayashi, Y. and Holleran, W. M. (2002) Glucosylceramide synthesis and synthase expression protect against ceramide-induced stress J. Lipid Res. 43, 1293-1302