フツウソバ(Fagopyrum esculentum)における生殖様式の遺伝的基盤の解明
植物の多くは、受粉時に自己の花粉と非自己の花粉を識別し、自己の花粉との受精を抑制する自他識別システムを有しています。この自他識別システムは自家不和合性とも呼ばれ、その情報ネットワーク制御の解明が急速に進んでいます。その中でも、Charles Darwinの著書「Different forms of flowers」(Darwin 1877)に掲載されているように、花の形態多型と自家不和合性が密接に関連した異形花型自家不和合性が知られています。異形花型自家不和合性は、他の植物でも見られる虫媒による他殖性強化のための巧妙な機構であり、花型と自家不和合性を統御する隣接遺伝子群がS座に集約されて機能していると考えられています(S-supergene; Barrett 2002)。
これまでに、私たちは、異形花型自家不和合性を示すフツウソバを材料として、S座に座乗する遺伝子S-ELF3を発見しました(Yasui, Aii ら 2012)。さらに、S-ELF3を失うと異形花型自家不和合性の表現型が、どのように変化するのかを調べるために、世界から収集されたフツウソバ遺伝資源の中から、S-ELF3を欠失した系統を探しました。そして、S-ELF3が欠失し、その周辺領域に変異が生じているS-del1を見出すに至りました。野生型のフツウソバは、雌しべと雄しべの長さが異なる花(短柱花と長柱花)を咲かせ、両者の間でしか受精が成立しません(図1)。ところが、S-del1は、雌しべと雄しべのの長さが等しい花(等長柱花)を咲かせ、自殖することができます。さらに、S-del1の花粉は、短柱花と長柱花に対しても受精することができます。
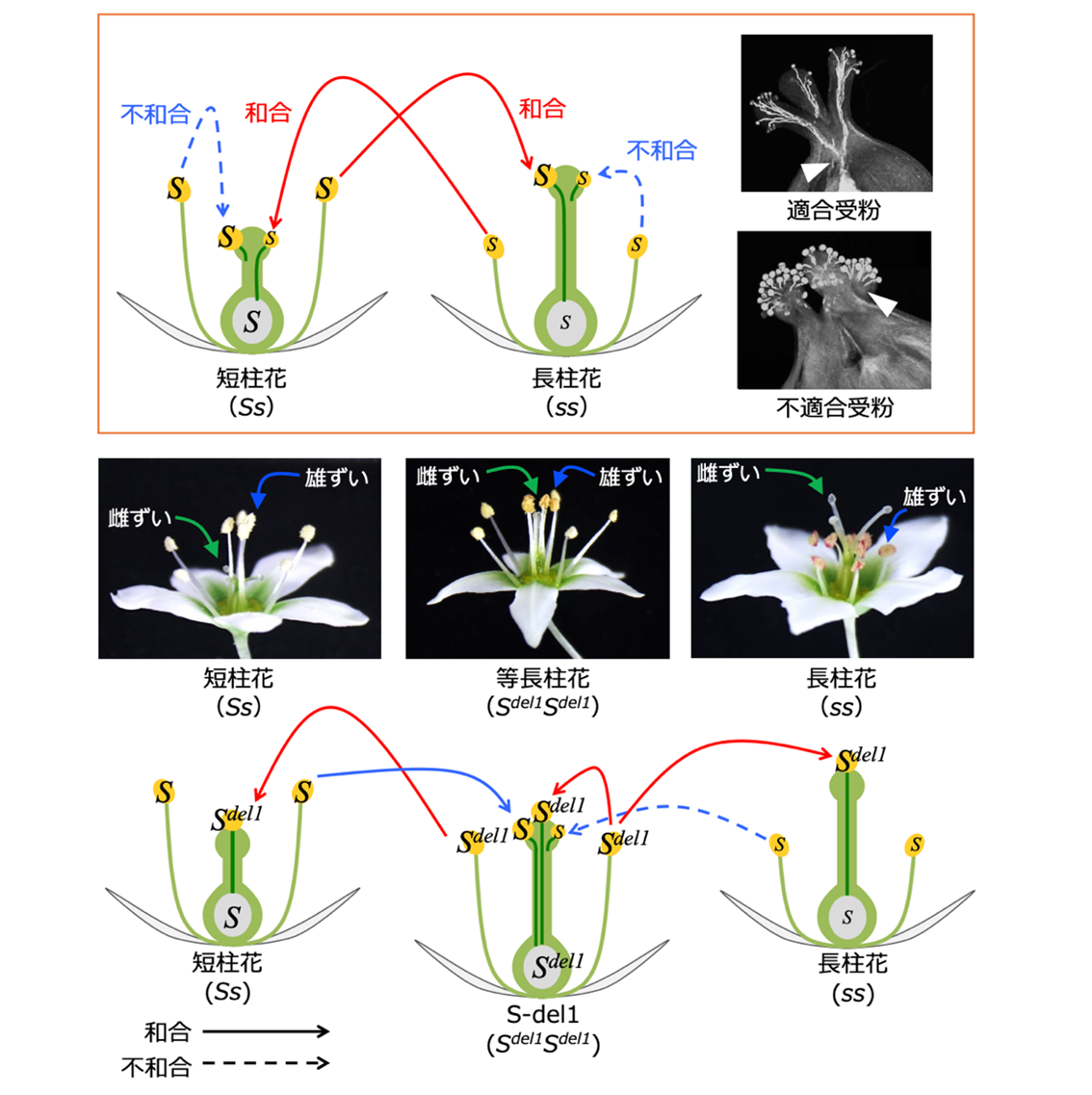
フツウソバは、雄しべが長く雌しべが短い花(短柱花)と、雄しべが短く雌しべが長い花(長柱花)をもつ個体が存在する異形花型自家不和合性植物です。短柱花のS座の遺伝型はSs、長柱花の遺伝型はssであり、両者間の交配により、後代の集団において両花型の存在が1:1に維持されます。私たちが発見した自家和合性変異体S-del1は、雄しべと雌しべの長さが等しい花(等長柱花)を咲かせます。S-del1のS座の遺伝型は、Sdel1Sdel1です。S-del1の花粉は、短柱花個体及び長柱花個体と受精することができます。
最近、Fawcett ら(2023)によって、S-ELF3は、フツウソバの異形花型自不和合性における雌性側の形質を制御する遺伝子であることが証明されました(図 2)。このことから、S-del1は、S-ELF3を失うことで、等長柱花の花を咲かせ自殖することができるようになったと考えられます。しかし、S-del1の花粉が「なぜ短柱花と受精できるのか」は不明です。

フツウソバのS座は、Sハプロタイプがヘミザイガスの状態であり、sハプロタイプの当該領域は欠失しています。自家和合性変異体S-del1のS座は、S-ELF3が座乗する周辺領域が欠失したSdel1ハプロタイプをもちます。フツウソバS座の雌性側因子は、S-ELF3であることがFawcettら(2023)によって証明されました。しかしながら、雄性側因子の実体は、未だ明らかになっていません。
この不思議な現象の理由を明らかにするために、私たちはS-del1を用いたゲノム科学・分子遺伝学的な解析を進めています。そして、これまでに得られた成果を統合することにより、フツウソバにおける異形花型自家不和合性の遺伝的基盤の解明を目指しています。